Microbiota-immune interactions: from gut to brain
Abstract
The vast diversity of bacteria that inhabit the gastrointestinal tract strongly influence host physiology, not only nutrient metabolism but also immune system development and function. The complexity of the microbiota is matched by the complexity of the host immune system, where they have coevolved to maintain homeostasis ensuring the mutualistic host-microbial relationship. Numerous studies in recent years investigating the gut-brain axis have demonstrated an important role for the gut microbiota in modulating brain development and function, with the immune system serving as an important coordinator of these interactions. Gut bacteria can modulate not only gut-resident immune cells but also brain-resident immune cells. Activation of the immune system in the gut and in the brain are implicated in responses to neuroinflammation, brain injury, as well as changes in neurogenesis and plasticity. Impairments in this bidirectional communication are implicated in the etiopathogenesis of psychiatric and neurodevelopmental diseases and disorders, including autism spectrum disorders, or comorbidities associated with Gastrointestinal diseases, including inflammatory bowel diseases, where dysbiosis is commonly seen. Consequently, probiotics, or beneficial microbes, are being recognized as promising therapeutic targets to modulate behavior and brain development by modulating the gut microbiota. Here we review the role of microbiota-immune interactions in the gut and the brain during homeostasis and disease and their impact on gut-brain communication, brain function, and behavior as well as the use of probiotics in central nervous system alterations.
Statement of novelty: The microbiota-gut-brain axis is increasingly recognized as an important physiological pathway for maintaining health and impacting the brain and central nervous system. Increasing evidence suggests that the immune system is crucial for gut-brain signaling. In this review, we highlight the critical studies in the literature that identify the key immune pathways involved.
Introduction
The intestine is the largest surface of the body and is constantly exposed to dietary and bacterial antigens as well as potentially noxious substances and infectious agents, which threaten the balance between health and disease (Salvo Romero et al. 2015). In mammals, the gastrointestinal (GI) tract harbors a complex microbial community, known as the intestinal microbiota, composed of thousands of different species of bacteria, viruses, fungi, archaea, and protists (Parfrey et al. 2011; Hillman et al. 2017). This complex community plays a critical role in the digestive process, protection against colonization with pathogens (Sonnenburg et al. 2004), as well as influencing the maturation and function of the intestinal immune system (Caricilli et al. 2014). In addition, there is increasing evidence that these microbial communities can regulate brain development, mood, and cognitive function through the bidirectional signaling between the gut and the brain (Deverman and Patterson 2009; Lakhan and Kirchgessner 2010). The complexity of the microbiota is matched by the complexity of the host immune system to ensure the maintenance of this balance and for preventing access of organisms to the host inner milieu. Chronic inflammation not only changes the microbiota composition, but it can also trigger significant and long-lasting behavioral changes, such as the development of cognitive impairment and depression, in a bottom-up manner (Lakhan and Kirchgessner 2010). Taken together, this suggests a critical role for the gut microbiota in regulating brain development and behavior, with the immune system emerging as an important coordinator of these interactions.
Despite the recent discovery of meningeal lymphatics reshaping our understanding of immunity within the central nervous system (CNS) (Louveau et al. 2015), the brain parenchyma is still considered “immune privileged” compared to peripheral tissues. The brain contains numerous resident immune cells that play a role in the defense against infection and injury, critically supporting neurons, and in maintaining and remodeling circuit connectivity, and regulating plasticity (Tremblay et al. 2011). Pathways that have been traditionally recognized for their functions in maintaining peripheral immunity are now known to be important in regulating neurodevelopment, including cytokines present in the developing brain, which can regulate neuronal differentiation, axonal growing, and synaptic plasticity (Deverman and Patterson 2009). In addition, expression of neuroendocrine receptors and factors on immune cells, such as neuropeptide receptors [i.e., calcitonin gene-related peptide (CGRP), substance (SP), adrenomedullin, neurokinins A and B, vasoactive intestinal peptide (VIP), neuropeptide Y (NPY), and gastrin releasing peptide (GRP), etc.], glucocorticoids, or adrenoreceptors can lead to the release of immune-derived mediators and are increasingly recognized for their role in the immune system (Hadden et al. 1970; Pert et al. 1985; Procaccini et al. 2014).
Microbial colonization of the GI tract has a significant impact on neurophysiology and behavior (Sampson and Mazmanian 2015). Given the immunomodulatory properties of the gut microbiota, peripheral immune cell pathways have been implicated as important mechanisms mediating microbial modulation of brain function and behavior (Rook et al. 2014; Rea et al. 2016), including neurodevelopmental disorders, such as autism spectrum disorder (ASD), and GI disease, such as inflammatory bowel disease (IBD), where psychosocial deficits commonly occur in patients. In this review, we discuss the roles for the gut microbiota as an integral mediator of neuroimmune interactions, examining how microbiota interacts between gut and brain-resident immune cells, and how they can impact the etiopathogenesis or manifestation of symptoms relevant to neurobehavioral and neurodegenerative disorders. Finally, we summarize effective probiotic interventions on the function of the CNS and discuss how probiotics can modulate this interaction.
The gut-brain axis
The gut-brain axis plays an important role not only in the proper maintenance of the GI tract but also brain function and behavior. This bidirectional signaling pathway is important in maintaining homeostasis and is regulated at different levels by the CNS, the enteric nervous system (ENS), the hypothalamic pituitary adrenal (HPA) axis, and immune system through complex communication by endocrine, immune, and neuronal factors (Figure 1) (Romijn et al. 2008; Weltens et al. 2018). CNS-mediated regulation of intestinal function and immune responses occurs through systemic, regional, and local pathways. The autonomic nervous system (ANS; sympathetic and parasympathetic branches; drives both afferent and efferent signals), the peripheral nervous system (through the release of neuropeptides), and the HPA axis (through the systemic release of glucocorticoids) are all involved. Moreover, neurally-mediated responses triggered by the activation of sensory afferents in response to changes in luminal content or pressure, muscle distention, and inflammation can be transmitted to target cells through reflexes of local enteric nerves, independent of the CNS (Carabotti et al. 2015).
Figure 1:
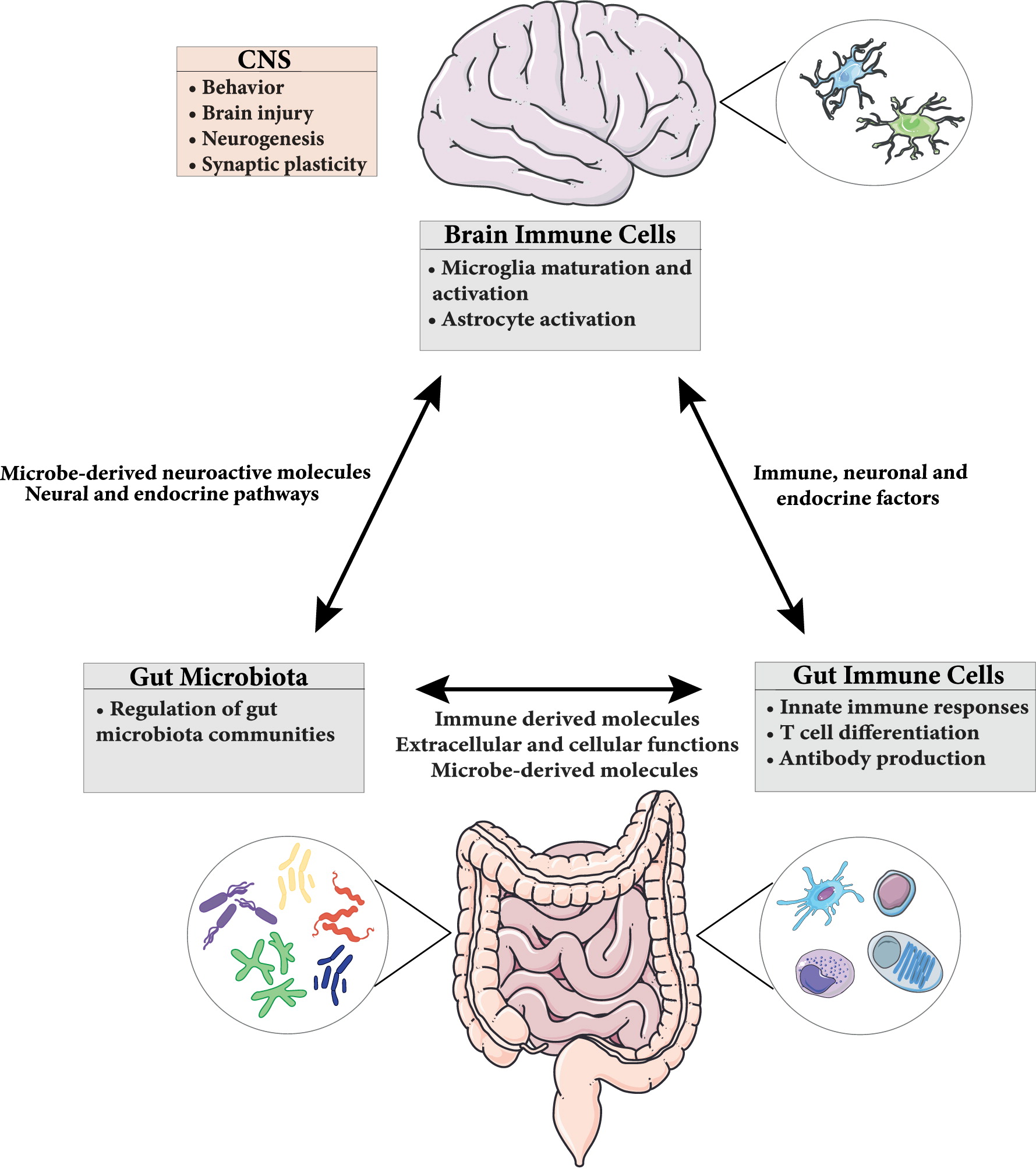
In the GI tract, in addition to the well-characterized intestinal epithelial cells (Sharkey and Mawe 2002) and resident immune cells (Dantzer 2018), the microbiota also critically contributes to brain-gut communication. This microbiota-gut-brain axis ensures not only the proper maintenance of GI function, but also maintains cognition and emotional behaviors (Martin and Mayer 2017). Moreover, disturbances of this system have been implicated in a wide range of disorders, including functional and inflammatory GI disorders such as irritable bowel syndrome (IBS) and IBD (Rhee et al. 2009; Mayer et al. 2014) as well as neurodevelopmental disorders, such as ASD (van De Sande et al. 2014).
Physical and biochemical barriers in the GI tract anatomically prevent the microbiota from entering the body and the underlying tissues, restricting it to the lumen. In case of a breach in barriers, immune cells in the GI tract are present to contain and restrict the passage of microbial components, while limiting inappropriate immune activation and overt inflammation (Hooper et al. 2012), which can impact the microbiota-gut-brain axis.
Microbiota-immune interactions in the gut
Impact of microbiota on immune system
The immune system is composed of a complex network of innate and adaptive cells and components endowed with an extraordinary capacity to respond to highly diverse challenges. Collectively, this cellular network acts as a regulator of host homeostasis to sustain and restore tissue function in the context of microbial and environmental encounters in the body. This homeostasis is crucial, with commensal microorganisms being required for the proper maturation of the immune system (Chung et al. 2012). This fundamental role of the microbiota has been derived from studies on germ-free (GF) mice or antibiotic-treated animals, supporting the concept that the development of the immune system coincides with the acquisition of a complex microbiota (Sharkey and Mawe 2002). GF mice exhibit immature gut-associated lymphoid tissues (GALT), decreased number of intestinal lymphocytes, decreased Immunoglobulin (Ig) A production and impaired antimicrobial peptides compared to colonized controls (Bauer et al. 1963; Hamada et al. 2002; Mazmanian et al. 2005; Smith et al. 2007). Many of these impairments can be restored by colonization of the microbiota, however some of these components can only be restored if colonization is performed early in development (Hansen et al. 2012).
The initial exposure of the immune system to commensal microbes occurs during birth via passage through the birth canal, and subsequently by maternal milk. Controversial evidence has suggested the potential for in utero exposure to microbes, however, this is likely limited to microbial metabolites or bacterial DNA, given the absence of viable microbes and the likelihood of contamination of placental samples during birth (Perez-Muñoz et al. 2017; Martinez et al. 2018; Theis et al. 2019). These early interactions are important to set the tone of the mucosal and systemic immune system into adulthood (Hooper et al. 2012). This capacity to establish tolerance of the immune system at an early age could be explained by the immaturity of the neonatal immune system, and the tolerogenic environment during development. For example, early in life the immune system has blunted inflammatory cytokine production, skewing T and B cell development to a more regulatory phenotype (Siegrist 2001; PrabhuDas et al. 2011). Toll-like receptors (TLRs), innate immune pattern recognition receptors, are involved in this differential recognition, being responsible for the normal development of the intestinal mucosal immune system which recognize microbial pathogens relevant to infants, such as group B Streptococcus, Listeria monocytogenes, and Respiratory Syncytial Virus (RSV) (Levy 2007). TLR-mediated cytokine production by mononuclear cells in vitro demonstrated less interferon (IFN)-α, IFN-γ, and interleukin (IL)-12 subunit p70 (IL-12-p70) expression from cells obtained from infants versus cells obtained from adults, which increases between birth and 1–2 years of age (PrabhuDas et al. 2011). Furthermore, TLR activation by antigens belonging to the normal intestinal microbiota signal the inhibition of inflammatory reactions and maintain epithelial barrier and protection from direct epithelial injury, suggesting an essential role of maintaining intestinal homeostasis (Rakoff-Nahoum et al. 2004). Complementarily, nucleotide-oligomerization-domain (Nod)-like receptors (NLRs) recognize various microbial specific molecules and trigger the assembly of inflammasomes, which can act as sensors of microbial-associated molecular patterns (MAMPs), with NLR deficiency associated with an altered immune response (Elinav et al. 2011) and impairments in behavior (Pusceddu et al. 2019). Microbiota colonization in the GI tract during early life affects the development of T cell populations into different types of T helper cells (Th) including: Th1, Th2, and Th17 or into regulatory T cells (Tregs) (Mazmanian et al. 2005; Gaboriau-Routhiau et al. 2009; Atarashi et al. 2011). For example, GF mice show reduced Treg cell induction, absence of Th17 cells, and an imbalance of Th1/Th2 ratio exhibiting a bias toward Th2 responses (Ivanov et al. 2009; Belkaid and Hand 2014). Colonization with Bacteroides fragilis recovered the development of the Th1-associated immune response through the bacterial product polysaccharide A-dependent pathway (Mazmanian et al. 2005). Moreover, B. fragilis also induced Treg accumulation through TLR-2 and inhibition of Th17 development (Round et al. 2011).
Antibody responses, while highly influenced by the microbiota, can still occur in GF animals (Zeng et al. 2018). Production of IgA by intestinal plasma cells is rapidly induced following microbial colonization and can occur through both T-dependent and T-independent pathways (Benckert et al. 2011; Bunker et al. 2015), which generates polyreactive IgA with low affinity to commensal bacteria, providing beneficial effects on surface-coated microbes. IgA is transcytosed and secreted through the epithelial layer into the lumen, and it has been shown that IgA coating can promote intestinal colonization of some commensals (Donaldson et al. 2018). Microbiota can modulate antibody production by generation of ATP through a mechanism that includes P2X7-mediated signaling of T follicular helper cells, which also limits the generation of protective IgA against enteropathogens (Proietti et al. 2014, 2019).
Impact of innate immune system on microbiota
In addition to the important role for rapid elimination of pathogens, innate immune responses also contribute to the establishment of the symbiotic relationship of the host with the microbiota. The intestinal innate immune system can also help define the overall niche of gut microbial communities by regulating secretion of defense mechanisms including mucus, antimicrobial peptides, and antibodies along the intestinal epithelial barrier (Kinnebrew and Pamer 2012).
The relationship between host immunity and the microbiota has been extensively explored using loss-of-function animal models (i.e., knockout [KO] mice). By using a simple model organism with an extremely simple gut commensal microbiota, Ryu et al. (2008) demonstrated that the immune status of the host can influence the composition of the microbiota, which can consequently dramatically impact the host. Using Drosophila melanogaster, defective regulation of nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB) signaling in the intestinal epithelium caused the overgrowth of a pathogenic commensal microbial community, leading to the death of the host (Ryu et al. 2008). However, in mammals, the enormous diversity of the resident microbiota community and the genetic complexity of the host immune system make it difficult to clearly establish the molecular links that would identify the unique interactions between immune genotype and commensal microbiota structure.
Several studies in mice also highlight the importance of innate immune responses for maintaining the mutualism for host-microbe interactions. In MyD88-deficient mice, defective in TLR signaling, intestinal dysbiosis was identified, predisposing mice to intestinal inflammation (Araki et al. 2005). Moreover, similar effects were shown in TLR-5 and NOD1/NOD2-double deficient mice, where alterations in microbiota composition were identified compared to WT mice (Kobayashi et al. 2005; Bouskra et al. 2008; Vijay-Kumar et al. 2010). Similarly, MyD88-mediated signaling in non-hematopoietic cells is required for clearance of the Gram positive intracellular pathogen L. monocytogenes from the GI tract by the antimicrobial C-type lectin Regenerating islet-derived protein 3 gamma (RegIIIγ) (Brandl et al. 2007). Humans and mice deficient in TLR/MyD88 signaling pathways also have diminished antimicrobial protein and mucus production (Vaishnava et al. 2008; Slack et al. 2009), however, translocation of bacteria into the blood stream is minimized by the compensation of high antibody titers. These antibody responses represent an important interaction between the 2 systems to regulate microbial-host responses in response to innate immune deficiency. Despite these initial studies, the lack of adequate littermate controls to standardize the microbiota of WT and KO groups largely prevented the ability to identify clear mechanistic insight into genetic regulation of the microbiota (Robertson et al. 2018). In this regard, other studies showed a little influence on the microbiota composition structure at homeostasis in KO mice defective in innate immune receptors (Robertson et al. 2018). For example, in Nod2−/− animals, where an impaired antibacterial response may influence the microbiota, genotype-dependent influences on the microbiota or differences in α-defensin secretions compared to wild-type (WT) controls were not observed (Robertson et al. 2013; Shanahan et al. 2014; Goethel et al. 2019). In addition, inflammasome-deficient mice, including Nlrp6-deficient mice and ASC-deficient mice which were initially thought to influence microbiota composition, showed no impact when the appropriate littermate controls were employed, highlighting the necessity of adequate littermate controls (Mamantopoulos et al. 2017). In this study, gut microbiota phylogenetic analyses were performed on ex-GF and littermate-controlled mice that were all allowed to shape their gut microbiota under physiological conditions after birth, minimizing non-genetic confounders (Mamantopoulos et al. 2017).
Despite the lack of an effect of innate immune impairment on the microbiota at homeostasis in Nod2−/− mice (Goethel et al. 2019), induction of inflammation and dysbiosis using antibiotics impaired microbial resilience in these mice. Similarly, neonatal antibiotic administration in Nod2−/− mice resulted in increased severity of colitis after administration of dextran sodium sulfate (DSS) to induce colitis (Goethel et al. 2019). During this acute inflammation in mice, the gut microbiota community shifts to resemble IBD-associated dysbiosis (Robertson et al. 2013), specifically with depletion of the beneficial short chain fatty acid (SCFA) butyrate-producing members of the microbiota reported (Rivera-Chavez et al. 2016). Taken together gene-deficiency in innate immune receptors appears to have little influence on microbiota structure in homeostasis, suggesting a redundancy in these gene functions and compensation of the adaptive system with respect to colonization of the microbiota. Conversely, their ability to influence bacterial structure occurs after the establishment of inflammation following an insult has been clearly established (Schwab et al. 2014).
Impact of adaptative immune system on the microbiota
Efficient immune responses in the GI tract are generated in highly organized structures, namely gut-associated lymphoid tissues (GALT). These secondary immune structures are only found in birds and mammals, not only for protection against infectious insults but also to allow and maintain complex bacterial communities in the gut. In addition, the microbiota of mammals appears to display evidence of coevolution, independently of the geographical location or diet (Ley et al. 2008a, 2008b). In mice, defects in the acquired immune system appear to have considerable effects on the microbiota. For example, Rag1−/− mice, which lack both T and B cells, have significantly decreased bacterial diversity than their littermate controls (Kwon et al. 2015). Similarly, mice lacking T-bet (T-box transcription factor family that regulates the development of the immune system) have severe intestinal inflammation and the promotion of an outgrowth of colitogenic bacteria (Garrett et al. 2007).
B cells are key players in mucosal immunity, with impairments in their function significantly impacting the intestinal microbiota. B-cell deficient mice have higher lipopolysaccharide (LPS) concentrations compared to WT control mice, coupled with colonization of fewer Clostricidiaceae and increased Paracoccus and Lactococcus genus, suggesting that antibody deficiency might change the species distribution within the microbial community (Shulzhenko et al. 2011). In the gut, most B cells differentiate into plasma cells, specifically IgA producing cells. The absence of IgA, or the impaired IgA selection in germinal centers due to deregulated T cell control, results in massive activation of the whole body immune system (Kawamoto et al. 2012; Nakajima et al. 2018). Interestingly, IgA promotes homeostatic bacterial composition by altering the expression of polysaccharide utilization loci (PUL), including a functionally uncharacterized molecular family provisionally named Mucus-Associated Functional Factor (MAFF), found in mice and humans, detected in mucus-resident bacteria, with expression requiring the presence of a complex microbiota (Nakajima et al. 2018).
T cells are also important in maintaining mucosal and systemic immunity. Tregs contribute to diversification of the gut microbiota, particularly by promoting species belonging to the Firmicutes phylum, by suppressing inflammation and regulation of IgA selection in Peyer’s patches (Kawamoto et al. 2014). Other adaptive immune cells such as IL-17 and IL-22-producing Th17 cells, can directly shape the microbial composition by promoting colonization of certain microbes and neutralization of invasive pathogens (Dubin and Kolls 2008; Bunker and Bendelac 2018). IL-22 deficiency or disruption of IL-17R signaling can severely affect the balance of gut bacterial communities, resulting in dysbiosis and increased susceptibility to intestinal inflammation (Ni et al. 2017).
Thus, the adaptive immune system, through cellular and molecular components that are required for immune tolerance and selection of antibody repertoire, help mediate host-microbial symbiosis, controlling the richness and balance of bacterial structures. Taken together, the immune system clearly exerts a central role in shaping the composition of the microbiota, while conversely, resident microbes provide signals that promote normal immune system development and influence immune responses. Disruption of these complex and dynamic interactions can have profound consequences for host health.
Microbiota-immune interactions in the brain
The maturation and development of the CNS is regulated by both intrinsic and extrinsic factors. Studies from GF mice, or those treated with broad-spectrum antibiotics, show that specific microbiota can impact CNS physiology and neurochemistry (Smith 2015), as they exhibit both neurological and cognitive deficits (Gareau et al. 2011), and neurotransmitter alterations compared to colonized controls (Strandwitz 2018). This communication between gut microbiota and the CNS is thought to be mediated by a combination of immune, enteric, and neural pathways providing a physical and chemical connection through the functional lymphatic vasculature and the blood-brain barrier (BBB). This suggests an important role for immune cells in CNS homeostasis as well as during microbial and environmental challenges (Louveau et al. 2015; Yoo and Mazmanian 2017).
Immune regulation within the CNS is mediated by microglia, astrocytes, and oligodendrocytes, given that the BBB limits access of circulating T and B cells to the underlying parenchyma. It has been demonstrated that the gut microbiota can influence and modulate microglial maturation and function (Erny et al. 2015), highlighting its critical role within the CNS (Braniste et al. 2014).
Microglia
Microglia are the tissue resident macrophages of the brain responsible for maintaining homeostasis, regulating synaptic transmission, synaptic pruning, and neuronal circuit formation (Schafer and Stevens 2015; Hong et al. 2016). Microglia work to protect the brain against various pathological conditions through immune activation, phagocytosis, and cytokine production (Nayak et al. 2014; Prinz et al. 2017). In GF mice, increased numbers of immature microglia across multiple brain areas such as the hippocampus, cerebellum, and cortex, are found compared to specific pathogen free (SPF)-colonized controls (Bercik et al. 2011a). These microglia show a more branching phenotype and downregulation in expression of genes that are associated with maturation to an active phenotype, highlighting their immaturity. Similarly, antibiotic treatment in colonized WT mice is associated with increased immature microglia, though total microglia numbers remain unchanged, with a normal phenotype restored following reintroduction of a complex microbiota (Erny et al. 2015). This suggests a role for the microbiota in microglia development that depends on developmental timing and (or) duration of microbial colonization. While the mechanisms of gut microbes influence on brain microglia remain unclear, they appear to be regulated by a great diversity and complexity of bacterial load more than by specific bacterial taxa. For example, GF mice colonized with a minimal community of bacterial species (Bacteroides distasonis, Lactobacillus salivarius and a member of Clostridium cluster XIV) maintained abnormalities in microglia (Schafer and Stevens 2015). However, the oral application of a mixture of the 3 major SCFAs acetate, propionate, and butyrate, was sufficient to drive maturation of microglia. It is known that SCFAs are able to cross the BBB and may therefore affect microglia directly (Huuskonen et al. 2004; Frost et al. 2014). While the mechanism of action is not well understood, SCFAs can act as signaling molecules by binding to G protein-coupled receptors, Gpr41 and Gpr43, which are highly expressed on intestinal epithelial and immune cells (Brown et al. 2003). Their activation can provoke a pro-inflammatory immune response, increase GI permeability, and increase absorption of neuro-active metabolites (Milo et al. 2002; Kim et al. 2013). SCFAs can also directly activate the sympathetic nervous system through Gpr41 that are found on sympathetic ganglionic neurons (Kimura et al. 2011), which can modulate catecholaminergic systems by affecting tyrosine hydroxylase gene transcription (DeCastro et al. 2005).
Immature microglia from GF mice are also impaired in their ability to respond to a stimulus. Microglia from GF mice display functionally impaired responses following challenge with LPS or lymphocytic choriomeningitis virus (LCMV), exhibiting altered morphology and attenuation of several genes relevant to interferon responses, pro-inflammatory cytokines, and effector processes. This suggests that gut immune-stimulatory products can influence microglia function to prevent CNS damage following viral or bacterial infection (Matcovitch-Natan et al. 2016).
Astrocytes
Astrocytes are the most abundant glial cell population in the brain and, similarly to microglia, have multiple functions in CNS integrity and homeostasis, including BBB integrity, ion gradient balance, neurotransmitter turnover and nutrient transport. They can also integrate information from adjacent glial, neuronal, vascular and immune cells to regulate neural excitability and synapse formation (Jensen et al. 2013; Khakh and Sofroniew 2015). Although they are not classically considered to be a CNS-resident immune cell, they (i) express MAMPs and major histocompatibility complex (MHC) class II antigen presenting receptor, (ii) are able to detect danger signals and respond via secretion of cytokines and chemokines, and (iii) activate adaptive immune defenses (Dong and Benveniste 2001; Farina et al. 2007).
The gut microbiota modulates astrocyte activity via microbial metabolites that activate aryl hydrocarbon receptors (AHR) (Rothhammer et al. 2016), a ligand-activated transcription factor that integrates environmental, dietary, microbial and metabolic cues to control complex transcriptional programs (Rothhammer and Quintana 2019). Gut microbiota-derived metabolites can bind to AHR and induce anti-inflammatory activity by restricting the recruitment of neurotoxic immune cells though type 1 IFN signaling. Studies with mice treated with the antibiotic ampicillin exhibit reduced AHR activation and decreased adverse disease symptoms. Ampicillin-sensitive bacteria can catalyze the conversion of dietary tryptophan to AHR agonists, and mice treated with ampicillin have decreased levels of AHR activation and worse disease scores in an experimental autoimmune encephalomyelitis (EAE) model, which is a commonly used mouse model of multiple sclerosis (MS). EAE mice supplemented with tryptophan have a reduction in pro-inflammatory molecules expressed in astrocytes and a reduction in symptom severity (Gareau et al. 2011; Strandwitz 2018). Microbial-derived tryptophan metabolites can also modulate astrocyte activation via microglia. Microglia activation by dietary tryptophan metabolites, through a mechanism also mediated by the AHR, modulate pro-inflammatory and neurotoxic activities in astrocytes in the EAE mouse model via production of TGFα and VEGF-B. Microglia-derived TGFα limit pathogenic activation via the ErbB1 receptor. Conversely, microglial VEGF-B triggers FLT-1 signaling in astrocytes and worsens EAE, including increasing demyelination and CNS monocyte recruitment (Rothhammer et al. 2018).
In human studies, astrocytes treated with tryptophan metabolites, including indole-3-proprionic acid and indole-3-aldehyde, have reduced expression of pro-inflammatory factors such as IL-6, IL-12, IL-23a, nitric oxide synthase 2 (Nos2) and CC chemokine family 2 (Ccl2), markers associated with MS in humans (Rothhammer et al. 2016). Taken together, these findings suggest that microbial metabolites of dietary tryptophan can modulate the inflammatory status of astrocytes thus contributing to neuroinflammation.
Microbiota-immune interactions and CNS alterations
Brain injury
Accumulating evidence indicates that brain-resident and peripheral immune cells are critically involved in MGB axis communication, with deficits in signaling contributing to inflammatory disorders and impaired neurogenesis in the CNS (Fung et al. 2017). Studies investigating the contributions of specific species of the microbiota to neuroinflammation have revealed complex interactions between intestinal bacteria, immune system activation, and neuroinflammation. For example, segmented filamentous bacteria (SFB) are epithelial-associated bacteria that promote the development of IL-17A-producing Th17 cells in the mouse small intestine (Gaboriau-Routhiau et al. 2009). Colonization of GF mice with SFB promotes the development of EAE symptoms via induction of Th17 cells (Lee et al. 2011). In EAE, brain autoimmunity is driven by a pro-inflammatory T cell response that promotes infiltration of immune cells in the CNS, with effector Th17 cells representing the first wave of pathogenic T cells (Korn et al. 2007) because of their ability to efficiently breach the blood-brain barrier (Kebir et al. 2007). On the other hand, colonization of mice with B. fragilis, which induces IL-10-producing regulatory Tregs, can reduce EAE severity. Additionally, metabolites produced by the gut microbiota, such as SCFAs, can also reduce EAE and axonal damage promoting Treg differentiation (Haghikia et al. 2015). In humans, excessive Th17 cell expansion in the intestine is associated with intestinal dysbiosis and brain autoimmunity. A higher Firmicutes/Bacteroidetes ratio, increased relative abundance of Streptococcus, and decreased Prevotella strains were observed in MS patients compared to healthy controls and with MS patients with no disease activity (Cosorich et al. 2017).
IL-17 and γδ T cells have also been implicated in ischemic stroke, a highly prevalent disease with limited therapeutic options, where inflammation is a key component in the pathophysiology (Iadecola and Anrather 2011). In a recent study, antibiotic-induced alterations in the intestinal microbiota were found to reduce ischemic brain injury in mice. This dysbiosis alters immune homeostasis through bacterial priming of intestinal dendritic cells, leading to an expansion of local Treg and a suppression of effector IL-17 and γδ T cells, thus improving outcome after brain ischemia (Benakis et al. 2016), and revealing an impact of the intestinal microbiota and meningeal IL-17 γδ T cells on ischemic injury.
Neurogenesis
Neurogenesis is an important physiological process that occurs throughout life and is critical for maintaining cognitive function (Aimone et al. 2014). Intestinal bacteria have recently been demonstrated to regulate both fetal and adult neurogenesis. For example, bacterial cell wall peptidoglycan (PGN), a universal MAMP for TLR2, traverses the maternal-fetal interface during maternal infection, leading to an altered neuroproliferative response and abnormal cognitive behavior during adulthood (Humann et al. 2016). In GF mice, studies assessing hippocampal neurogenesis reveal increased proliferation relative to conventional mice (Ogbonnaya et al. 2015). However, post-weaning exposure of GF mice to microbes could not influence neurogenesis, suggesting a critical time period where neuronal growth is stimulated by the microbiota during early post-natal development. In studies using antibiotic administration in adult animals for 7 weeks to reduce the microbiota abundance and diversity, decreases in hippocampal neurogenesis and spatial and object recognition were observed (Mohle et al. 2016). Reconstitution with normal SPF gut microbiota by fecal microbiota transplantation did not completely reverse the deficits in neurogenesis, unless the mice were supplemented with probiotics or exercise. Cells of the innate immune system, in particular Ly6Chi monocytes, were critical for the restorative effect (Mohle et al. 2016). This suggests that modification of the microbiota coupled with environmental enrichment (Noack et al. 2014) could help to restore monocyte homeostasis and brain plasticity and may help to counteract side effects of prolonged antibiotic treatment.
Synaptic plasticity and memory
Brain plasticity, a critical process involving change in structure and function of neurons following stimulation, is crucial for learning and memory. Brain plasticity results in multiple, dissociable changes including increases in dendritic length, alterations in spine density, synapse formation, increased glial activity, and altered metabolic activity (Kolb and Whishaw 1998). Cytokines, together with neurotransmitters and hormones, are signaling molecules playing a key role in the maintenance of neuro-immune-endocrine system homeostasis in the brain from early development until aging (Alboni and Maggi 2015). Early exposure to infectious agents or environmental stressors can influence reactivity to stress, immune regulation, and susceptibility to disease later in life. For instance, neonatal exposure to bacterial products (e.g., LPS) increases HPA responses to stress, decreases susceptibility to inflammation, and attenuates fever in response to a subsequent challenge in adult rats (Shanks et al. 2000; Boissé et al. 2004). Pro-inflammatory cytokines are important mediators of abnormal brain development during perinatal infection, and behavioral alterations following infection may occur as a result of an increase in pro-inflammatory cytokines, particularly in the hippocampus, where cytokine receptors are widely expressed (Cunningham and De Souza 1993; Cai et al. 2000; Urakubo et al. 2001). Immune cells are increasingly implicated for their role in synaptic plasticity mechanisms within the brain, including memory, particularly during early life.
Neonatal systemic exposure to non-pathogenic E. coli in rats causes memory impairments. This impairment, however, was only observed following a secondary peripheral immune challenge with LPS following contextual fear conditioning exposure, where increased astrocyte reactivity in the hippocampus was shown. The pro-inflammatory cytokine IL-1β is induced within the hippocampus in response to normal learning and is critical for maintaining long-term-potentiation (LTP) (Cunningham and De Souza 1993; Ross et al. 2003). Mice lacking IL-1β, or its type 1 receptor, exhibit markedly impaired hippocampal-dependent learning and memory (Goshen et al. 2007; Spulber et al. 2009). High levels of IL-1β can profoundly impair memory (Barrientos et al. 2002, 2009), with increased levels observed in patients with Alzheimer’s disease (AD) and other neurodegenerative disorders such as Parkinson’s disease and amyotrophic lateral sclerosis (Griffin et al. 1989; Stanley et al. 1994). In fact, peripherally-induced neuroinflammation modeling neurodegenerative disease are driven by LPS administration, which has become an important tool for deciphering pathological mechanisms involved in neurodegeneration (Catorce and Gevorkian 2016). In neonatally infected rats, a marked deficit in hippocampal-dependent memory in adulthood was observed, but similar to the previously mentioned studies, only after a subsequent immune challenge with LPS at the time of learning. Cluster of differentiation molecule 11b (CD11b)-enriched cells, a marker for microglia, produced exaggerated IL-1β ex vivo and after LPS exposure prior to learning in vivo, implicating the necessity of a second hit to impact memory dysfunction once a long-term change within the immune system has been stablished in early life. Prevention of microglial activation during learning prevents memory impairment in infected rats (Williamson et al. 2011). Taken together, these studies highlight a critical role for the immune system in mediating learning and memory.
Behavior
The host immune system is also critical in the development of behavioral and cognitive function. Consequently, long-term peripheral illnesses and chronic intestinal inflammation are associated with behavioral disturbances such as cognitive impairment, deficits in learning, impaired memory, depression and anxiety, which is linked to disrupted adult hippocampal neurogenesis in animal models (Zonis et al. 2015; Chesnokova et al. 2016).
ASD
ASD is a neurodevelopmental disorder characterized by stereotypic behavior, language deficits, and impaired social interaction. A high proportion of patients also suffer from GI disturbances (Hsiao 2014), including altered bowel habits in the context of intestinal dysbiosis, suggesting it represents a gut-brain disorder. In fact, GI disturbances have been specifically associated with changes in microbial communities in ASD patients (Brandl et al. 2007; Vuong and Hsiao 2017). In a pilot study, children diagnosed with ASD showed an improvement in both GI symptoms and aberrant behaviors following 5 weeks of administration of B. infantis (Sanctuary et al. 2019). Similarly in a mouse model, probiotic administration of Bacteroides fragilis resulted in restoration of gut permeability and improvement in some behavioral abnormalities (Hsiao et al. 2013). Social behavior, another common behavioral alteration in ASD, was detrimentally impacted in offspring following exposure to maternal high fat diet, and associated with a shift in intestinal microbial ecology (Buffington et al. 2016). Social deficits in offspring can also be rescued by administration of the probiotic L. reuteri (Buffington et al. 2016), which was found to act via the vagus nerve (Sgritta et al. 2019).
In maternal immune activation (MIA), a well-established animal model of ASD consisting of administration of the TLR3 agonist poly(I:C) in pregnant mice, offspring develop behavioral phenotypes similar to those observed in ASD, including anxiety, repetitive behavior and altered sociability (Hsiao et al. 2013). MIA offspring also exhibit GI mucosal barrier defects and intestinal dysbiosis compared to sham controls (Hsiao et al. 2013). Oral treatment with the human commensal B. fragilis restored intestinal permeability defects, as well as microbiota dysbiosis, and significantly improved behavioral abnormalities (Hsiao et al. 2013). IL-17A was implicated as a critical mediator inducing behavioral and cortical abnormalities in offspring in response to maternal intestinal bacteria that promotes Th17 differentiation. Pregnant mice colonized with the commensal SFB, or human commensal bacteria that induce intestinal Th17 cells, were more susceptible to having offspring with MIA-associated abnormalities (Kim et al. 2017), linking MIA-induced immune dysregulation to intestinal dysbiosis. Finally, in a genetic model of ASD using inbred BTBR T+Itpr3tf/J mouse (BTBR) mice, authors showed a reduction in the relative abundance of a bile-metabolizing Bifidobacterium and Blautia species, a particular bacterial taxa, associated with impaired social interactions as well as GI dysfunction and bile acid and altered tryptophan metabolism in the intestine (Golubeva et al. 2017). Together these studies support the concept of microbiota-based interventions on ASD behavior.
IBD
GI diseases are associated with a high risk for extra-intestinal manifestations, particularly psychiatric disturbance, which significantly impact patient quality of life. IBDs, consisting of Crohn’s disease (CD) and ulcerative colitis (UC), have a strong association with behavioral abnormalities in patients suffering from both active or resolved GI inflammation (van Langenberg et al. 2017). Adolescent IBD patients are particularly vulnerable to behavioral comorbidities and have been demonstrated to display mild cognitive problems, particularly with verbal memory (Filipovic and Filipovic 2014; van Langenberg et al. 2017), thought to be related to the significant brain development occurring during this critical developmental period. Stress significantly contributes to the development and (or) exacerbation of IBD (de Punder and Pruimboom 2015). Few clinical studies have also revealed that stress is associated with digestive problems and poor GI health (Walker et al. 2001). For instance, maternal prenatal stress was strongly and persistently associated with aberrant microbial colonization patterns in pediatric patients and associated with increased infant GI symptoms and allergic reactions (Zijlmans et al. 2015). Additionally, stress-related disorders such as depression are also associated with increased bacterial translocation and increased immune responses against commensal microbiota (Maes et al. 2012).
The effect of the MGB axis and its implication in behavior in IBD has increasingly been studied using mouse models. In a chronic murine DSS model, anxiety-like behavior was demonstrated after 3 cycles of DSS-induced colonic inflammation, which could be ameliorated by administration of a Bifidobacterium-containing probiotic via a vagally mediated pathway (Bercik et al. 2011b). In acute inflammation, changes in mood and behavior, including anxiety-like behavior, were observed following DSS-induced colitis, which were associated with a shift in the composition of the gut microbiota due to colonic inflammation (Emge et al. 2016). Similarly to the chronic DSS study, administration of probiotics could normalize changes in behavior, which widely correlated with restoration of the microbiota (Emge et al. 2016).
Probiotics on central nervous system functions
Probiotics are defined as “live micro-organisms which, when administered in adequate amounts, confer a health benefit on the host” (Morelli and Capurso 2012). While their exact mechanisms of action remain to be completely elucidated, probiotics are considered a novel and relatively safe way (Yelin et al. 2019) to beneficially modulate the gut microbiota for many GI and extra-intestinal diseases.
Multiple studies have used probiotics to modify the MGB axis, with improvements in behaviors associated with stress-related psychiatric conditions, improved memory abilities, and induction of neuronal plasticity. For instance, probiotic-induced promotion of neurogenesis by increasing hippocampal brain derived neurotropic factor (BDNF) expression normalizes the abnormal response of the HPA axis in mice (Ait-Belgnaoui et al. 2014). B. longum, B. breve, B. infantis, L. helveticus, L. rhamnosus, L. plantarum, and L. casei were the most commonly used probiotics for beneficially impacting behaviors, as either single- or multi-strain preparations, all of which were able to improve anxiety, depression, and memory related behaviors, based on multiple animal models (Wang et al. 2016). For example in depression, each of the single strains of B. longum, B. breve, L. rhamnosus, and L. helveticus all showed antidepressant effects (Desbonnet et al. 2008; Bravo et al. 2011; Singh et al. 2012; Ohland et al. 2013). In another study in both rats and mice, these same strains also improved spatial and non-spatial memory abilities (Luo et al. 2014; Liang et al. 2015; Savignac et al. 2015).
In humans, many studies demonstrated a beneficial effect of probiotics on psychiatric conditions in both patients and healthy individuals, although not all studies found a benefit. With respect to anxiety and depression, a probiotic formulation of B. longum and L. helveticus could improve anxiety and depression in all participants (Messaoudi et al. 2011a, 2011b). Other probiotic strains also have effects on anxiety and depression. A probiotic yogurt containing B. lactis and L. acidophilus improved general health questionnaire (GHQ) and depression anxiety and stress scale (DASS) scores on petrochemical workers (Mohammadi et al. 2016) and a recent study using multi-strain probiotics found improvement in the Leiden index of depression sensitivity scale (LEIDS-r score) in healthy participants, demonstrated by a significantly reduced overall cognitive reactivity to sad mood, which is predictive of depression (Steenbergen et al. 2015).
Similarly, in a study in chronic fatigue syndrome patients, L. casei Shirota administration decreased anxiety levels following treatment (Rao et al. 2009; Ait-Belgnaoui et al. 2014). In contrast, patients with schizophrenia showed no changes in the Positive and Negative Symptom Scale Score (PANSS) after 14 weeks following L. rhamnosus administration (Dickerson et al. 2014). Similarly, patients with rheumatoid arthritis showed no change in anxiety levels after intervention with L. casei for 8 weeks (Vaghef-Mehrabany et al. 2014). A recent study in healthy volunteers using the Paired Associate Learning test from the Cambridge Neuropsychological Test showed improvements of visuospatial memory performance following intake of B. longum 1714 (Allen et al. 2016). Finally, human immunodeficiency virus patients treated with a multi-strain probiotic supplementation (Vivomixx®; Visbiome®) also demonstrated improved neurocognitive functions including verbal and visual memory related to memory improvement (Ceccarelli et al. 2017). Currently, only animal studies have provided strong evidence for a beneficial effect of probiotics on cognition and memory, therefore additional clinical studies are needed to adequately test these effects. The effect of probiotics on the CNS in humans is reviewed extensively elsewhere (Wang et al. 2016; Cryan et al. 2019).
Many of these probiotics presumably act beneficially in part by inhibiting the growth of other harmful bacteria or pathogens and (or) improving the immune system (Luo et al. 2014; Liang et al. 2015). The proposed mechanisms of actions by which probiotics have a beneficial effect are diverse and comprise endocrine, immune, neural, and metabolic pathways. Many probiotics have shown a reduction in HPA axis activity by decreasing corticosteroid and (or) adrenocorticotropic hormone (ACTH) levels. Our laboratory identified changes in neurogenesis and synaptic plasticity with effects on memory by modulating HPA axis activity. Using a combination of L. rhamnosus + L. helveticus, improved memory and cFOS expression, an immediate early gene serving as a neuronal activation marker, was found in mice infected with C. rodentium following exposure to an acute psychological stress (Gareau et al. 2011).
Probiotics can also influence the CNS directly via the vagus nerve and by modulating neurotransmitter levels. Treatments that target the vagus nerve increase the vagal tone and inhibit cytokine production. The stimulation of vagal afferent fibers in the gut influences monoaminergic brain systems within the brain stem that play crucial roles in major psychiatric conditions, such as mood and anxiety disorders. For example, behavioral changes following L. rhamnosus and B. longum administration, were not seen in vagotomized animals (Bercik et al. 2011b). Furthermore, B. fragilis could enhance serum tryptophan levels, a precursor of 5-hydroxytryptamine (5-HT) or serotonin synthesis (Emge et al. 2016). Serotonin is an important neurotransmitter in the gut that can stimulate peristalsis and induce nausea and vomiting by activating the vagus nerve. Interactions between the vagus nerve and serotonin systems, with the gut microbiota controlling tryptophan catabolism, appear to play an important role in the treatment of psychiatric conditions (Breit et al. 2018).
Finally, another mechanism of action of probiotics altering CNS function is thought to be by decreasing pro-inflammatory cytokines and increasing anti-inflammatory cytokines. Cultured intestinal mucosal tissues of CD patients with L. casei, L. bulgaricus, L. crispatus and E. coli showed an interaction with immunocompetent cells and modulation of the production of pro-inflammatory cytokines, resulting in a significant reduction in the pro-inflammatory cytokine TNF-α (Borruel et al. 2002). In IL-10-deficient mice, which spontaneously develop colitis, significant reductions of INF-γ and TNF-α by Peyer’s patch lymphocytes and pro-inflammatory cytokine production by splenocytes were found in probiotic-treated mice (McCarthy et al. 2003). In healthy rats, the administration of a mixture of Lactobacillus and Bifidobacterium upregulated IL-10 and downregulated TNF-α and IL-6 (Karamese et al. 2016). Moreover, probiotics can ameliorate inflammatory immune responses through modulation of the intestinal barrier permeability (Llopis et al. 2005, 2009; Rao and Samak 2013). Increased intestinal barrier permeability is associated with psychiatric disorders, such as depression and ASD, while it can be restored by probiotic formulations of B. longum and L. helveticus, along with improved CNS function (Arseneault-Breard et al. 2012; Hsiao et al. 2013).
It is important to highlight the limitations of probiotics studies in both animals and in humans. With respect to the translation of behavioral models in animals to human health, it is important to note that the tests used to measure behavior in animals have no direct equivalents in humans. Theses behavioral tests are designed to reflect the presumed CNS dysfunction in humans, based on specific animal behaviors. The common use of questionnaires in humans to assess behaviors could result in subjective biases. In the future, use of neuroimaging techniques may provide a better, more accurate, alternative way to assess changes in brain function (McFarland et al. 2018). Several studies have found that in contrast to inbred mice, humans present individual, regional, and strain-specific mucosal bacterial colonization patterns. In some individuals, these unique features prevents standard probiotics from transiently colonizing the gut, as determined by indistinguishable probiotic presence in the stool (Zmora et al. 2018). In the absence of individual-specific mucosal colonization to probiotics, lack of beneficial effects could be misunderstood (Llopis et al. 2009). Of importance for future studies will be the development of personalized medicine using strain-specific and individual-specific based probiotic therapy based on next-generation sequencing, to enable persistent live-bacteria colonization to beneficially impact the host.
Conclusions
Accumulating evidence indicates that brain-resident and gut-resident immune cells are critically involved in orchestrating MGB axis communication. The gut microbiota can influence the development and function of the intestinal immune system and conversely, the innate and adaptive immune system influence microbiota composition. This reciprocal relationship contributes to establishment of symbiosis with the gut microbiota, preventing potentially harmful bacteria and pathogens from breaching the host defenses. The microbiota can also modulate brain microglia and astrocytes, which mediate neurophysiological processes including neural development, neurotransmission, and CNS immune activation. These microbial influences on immune responses have important consequences for brain inflammation, injury, behavior and brain plasticity and are increasingly associated with symptoms of various neuroinflammatory and psychiatric disorders. This relationship establishes a tight communication with the CNS and immune system as important cellular mediators across the gut-brain axis. Disruption of these complex and dynamic interactions can have profound consequences for host health.
Studies of probiotics in animal models, and a few studies in humans, have identified their effective application in the prevention and treatment of various health conditions and diseases such as GI infections but also in improving CNS function, including psychiatric disorders such as anxiety and depression, and cognition.
Finally, as the specific pathways of the MGB axis signaling are still incompletely characterized, it is critical for future studies to clarify these pathways to define specific microbe-derived factors, immune effector functions, and microbiota–immune pathways for modulating brain function and behavior. Also, it is crucial for future microbiota studies that appropriate controls be used to compare baseline variation in experimental groups. Likewise, it is important to note that laboratory mice have distinct immunological profiles from humans, limiting the ability to directly extrapolate conclusions to human diseases. Future mechanistic insights into how the immune system and microbiota interact will require development of novel approaches with high throughput analysis of metabolites as well as gene and protein expression, to move towards a personalized-based therapy.
Acknowledgements
This research was supported by the NIH (1R01AT009365-01 and 5R21MH108154-01 to MGG).
REFERENCES
Aimone J.B., Li Y., Lee S.W., Clemenson G.D., Deng W., and Gage F.H. 2014. Regulation and function of adult neurogenesis: From genes to cognition. Physiol. Rev. 94:991–1026.
Ait-Belgnaoui A., Colom A., Braniste V., Ramalho L., Marrot A., Cartier C., Houdeau E., Theodorou V., and Tompkins T. 2014. Probiotic gut effect prevents the chronic psychological stress-induced brain activity abnormality in mice. Neurogastroenterol. Motil. 26:510–520.
Alboni S. and Maggi L. 2015. Editorial: Cytokines as players of neuronal plasticity and sensitivity to environment in healthy and pathological brain. Front. Cell. Neurosci. 9:508.
Allen A.P., Hutch W., Borre Y.E., Kennedy P.J., Temko A., Boylan G., Murphy E., Cryan J.F., Dinan T.G., and Clarke G. 2016. Bifidobacterium longum 1714 as a translational psychobiotic: Modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl. Psychiatry. 6:e939.
Araki A., Kanai T., Ishikura T., Makita S., Uraushihara K., Iiyama R., Totsuka T., Takeda K., Akira S., and Watanabe M. 2005. MyD88-deficient mice develop severe intestinal inflammation in dextran sodium sulfate colitis. J. Gastroenterol. 40:16–23.
Arseneault-Breard J., Rondeau I., Gilbert K., Girard S.A., Tompkins T.A., Godbout R., and Rousseau G. 2012. Combination of Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 reduces post-myocardial infarction depression symptoms and restores intestinal permeability in a rat model. Br. J. Nutr. 107:1793–1799.
Atarashi K., Tanoue T., Shima T., Imaoka A., Kuwahara T., Momose Y., Cheng G., Yamasaki S., Saito T., Ohba Y., Taniguchi T., Takeda K., Hori S., Ivanov I.I., Umesaki Y., Itoh K., and Honda K. 2011. Induction of colonic regulatory T cells by indigenous Clostridium species. Science. 331:337–341.
Barrientos R.M., Higgins E.A., Sprunger D.B., Watkins L.R., Rudy J.W., and Maier S.F. 2002. Memory for context is impaired by a post context exposure injection of interleukin-1 beta into dorsal hippocampus. Behav. Brain Res. 134:291–298.
Barrientos R.M., Frank M.G., Hein A.M., Higgins E.A., Watkins L.R., Rudy J.W., and Maier S.F. 2009. Time course of hippocampal IL-1 beta and memory consolidation impairments in aging rats following peripheral infection. Brain Behav. Immun. 23:46–54.
Bauer H., Horowitz R.E., Levenson S.M., and Popper H. 1963. The response of the lymphatic tissue to the microbial flora. Studies on germfree mice. Am. J. Pathol. 42:471–483.
Belkaid Y. and Hand T.W. 2014. Role of the microbiota in immunity and inflammation. Cell. 157:121–141.
Benakis C., Brea D., Caballero S., Faraco G., Moore J., Murphy M., Sita G., Racchumi G., Ling L., Pamer E.G., Iadecola C., and Anrather J. 2016. Commensal microbiota affects ischemic stroke outcome by regulating intestinal γδ T cells. Nat. Med. 22:516–523.
Benckert J., Schmolka N., Kreschel C., Zoller M.J., Sturm A., Wiedenmann B., and Wardemann H. 2011. The majority of intestinal IgA+ and IgG+ plasmablasts in the human gut are antigen-specific. J. Clin. Invest. 121:1946–1955.
Bercik P., Denou E., Collins J., Jackson W., Lu J., Jury J., Deng Y., Blennerhassett P., Macri J., McCoy K.D., Verdu E.F., and Collins S.M. 2011a. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology. 141:599–609.
Bercik P., Park A.J., Sinclair D., Khoshdel A., Lu J., Huang X., Deng Y., Blennerhassett P.A., Fahnestock M., Moine D., Berger B., Huizinga J.D., Kunze W., McLean P.G., Bergonzelli G.E., Collins S.M., and Verdu E.F. 2011b. The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroenterol. Motil. 23:1132–1139.
Boissé L., Mouihate A., Ellis S., and Pittman Q.J. 2004. Long-term alterations in neuroimmune responses after neonatal exposure to lipopolysaccharide. J. Neurosci. 24:4928–4934.
Borruel N., Carol M., Casellas F., Antolin M., De Lara F., Espin E., Naval J., Guarner F., and Malagelada J.R. 2002. Increased mucosal tumour necrosis factor alpha production in Crohn’s disease can be downregulated ex vivo by probiotic bacteria. Gut. 51:659–664.
Bouskra D., Brézillon C., Bérard M., Werts C., Varona R., Boneca I.G., and Eberl G. 2008. Lymphoid tissue genesis induced by commensals through NOD1 regulates intestinal homeostasis. Nature. 456:507–510.
Brandl K., Plitas G., Schnabl B., DeMatteo R.P., and Pamer E.G. 2007. MyD88-mediated signals induce the bactericidal lectin RegIII gamma and protect mice against intestinal Listeria monocytogenes infection. J. Exp. Med. 204:1891–1900.
Braniste V., Al-Asmakh M., Kowal C., Anuar F., Abbaspour A., Tóth M., Korecka A., Bakocevic N., Ng L.G., Kundu P., Gulyás B., Halldin C., Hultenby K., Nilsson H., Hebert H., Volpe B.T., Diamond B., and Pettersson S. 2014. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 6:263ra158.
Bravo J.A., Forsythe P., Chew M.V., Escaravage E., Savignac H.M., Dinan T.G., Bienenstock J., and Cryan J.F. 2011. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA. 108:16050–16055.
Breit S., Kupferberg A., Rogler G., and Hasler G. 2018. Vagus nerve as modulator of the brain-gut axis in psychiatric and inflammatory disorders. Front. Psychiatry. 9:44.
Brown A.J., Goldsworthy S.M., Barnes A.A., Eilert M.M., Tcheang L., Daniels D., Muir A.I., Wigglesworth M.J., Kinghorn I., Fraser N.J., Pike N.B., Strum J.C., Steplewski K.M., Murdock P.R., Holder J.C., Marshall F.H., Szekeres P.G., Wilson S., Ignar D.M., Foord S.M., Wise A., and Dowell S.J. 2003. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 278:11312–11319.
Buffington S.A., Di Prisco G.V., Auchtung T.A., Ajami N.J., Petrosino J.F., and Costa-Mattioli M. 2016. Microbial reconstitution reverses maternal diet-induced social and synaptic deficits in offspring. Cell. 165:1762–1775.
Bunker J.J. and Bendelac A. 2018. IgA responses to microbiota. Immunity. 49:211–224.
Bunker J.J., Flynn T.M., Koval J.C., Shaw D.G., Meisel M., McDonald B.D., Ishizuka I.E., Dent A.L., Wilson P.C., Jabri B., Antonopoulos D.A., and Bendelac A. 2015. Innate and adaptive humoral responses coat distinct commensal bacteria with immunoglobulin A. Immunity. 43:541–553.
Cai Z., Pan Z.L., Pang Y., Evans O.B., and Rhodes P.G. 2000. Cytokine induction in fetal rat brains and brain injury in neonatal rats after maternal lipopolysaccharide administration. Pediatr. Res. 47:64–72.
Carabotti M., Scirocco A., Maselli M.A., and Severi C. 2015. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 28:203–209.
Caricilli A.M., Castoldi A., and Camara N.O. 2014. Intestinal barrier: A gentlemen’s agreement between microbiota and immunity. World J. Gastrointest. Pathophysiol. 5:18–32.
Catorce M.N. and Gevorkian G. 2016. LPS-induced murine neuroinflammation model: Main features and suitability for pre-clinical assessment of nutraceuticals. Curr. Neuropharmacol. 14:155–164.
Ceccarelli G., Fratino M., Selvaggi C., Giustini N., Serafino S., Schietroma I., Corano Scheri G., Pavone P., Passavanti G., Alunni Fegatelli D., Mezzaroma I., Antonelli G., Vullo V., Scagnolari C., and d’Ettorre G. 2017. A pilot study on the effects of probiotic supplementation on neuropsychological performance and microRNA-29a-c levels in antiretroviral-treated HIV-1-infected patients. Brain Behav. 7:e00756.
Chesnokova V., Pechnick R.N., and Wawrowsky K. 2016. Chronic peripheral inflammation, hippocampal neurogenesis, and behavior. Brain Behav. Immun. 58:1–8.
Chung H., Pamp S.J., Hill J.A., Surana N.K., Edelman S.M., Troy E.B., Reading N.C., Villablanca E.J., Wang S., Mora J.R., Umesaki Y., Mathis D., Benoist C., Relman D.A., and Kasper D.L. 2012. Gut immune maturation depends on colonization with a host-specific microbiota. Cell. 149:1578–1593.
Cosorich I., Dalla-Costa G., Sorini C., Ferrarese R., Messina M.J., Dolpady J., Radice E., Mariani A., Testoni P.A., Canducci F., Comi G., Martinelli V., and Falcone M. 2017. High frequency of intestinal TH17 cells correlates with microbiota alterations and disease activity in multiple sclerosis. Sci. Adv. 3:e1700492.
Cryan J.F., O’Riordan K.J., Cowan C.S.M., Sandhu K.V., Bastiaanssen T.F.S., Boehme M., Codagnone M.G., Cussotto S., Fulling C., Golubeva A.V., Guzzetta K.E., Jaggar M., Long-Smith C.M., Lyte J.M., Martin J.A., Molinero-Perez A., Moloney G., Morelli E., Morillas E., O’Connor R., Cruz-Pereira J.S., Peterson V.L., Rea K., Ritz N.L., Sherwin E., Spichak S., Teichman E.M., van de Wouw M., Ventura-Silva A.P., Wallace-Fitzsimons S.E., Hyland N., Clarke G., and Dinan T.G. 2019. The microbiota-gut-brain axis. Physiol. Rev. 99:1877–2013.
Cunningham E.T. Jr. and De Souza E.B. 1993. Interleukin 1 receptors in the brain and endocrine tissues. Immunol. Today. 14:171–176.
Dantzer R. 2018. Neuroimmune interactions: From the brain to the immune system and vice versa. Physiol. Rev. 98:477–504.
de Punder K. and Pruimboom L. 2015. Stress induces endotoxemia and low-grade inflammation by increasing barrier permeability. Front. Immunol. 6:223.
DeCastro M., Nankova B.B., Shah P., Patel P., Mally P.V., Mishra R., and La Gamma E.F. 2005. Short chain fatty acids regulate tyrosine hydroxylase gene expression through a cAMP-dependent signaling pathway. Brain Res. Mol. Brain Res. 142:28–38.
Desbonnet L., Garrett L., Clarke G., Bienenstock J., and Dinan T.G. 2008. The probiotic Bifidobacteria infantis: An assessment of potential antidepressant properties in the rat. J. Psychiatr. Res. 43:164–174.
Deverman B.E. and Patterson P.H. 2009. Cytokines and CNS development. Neuron. 64:61–78.
Dickerson F.B., Stallings C., Origoni A., Katsafanas E., Savage C.L., Schweinfurth L.A., Goga J., Khushalani S., and Yolken R.H. 2014. Effect of probiotic supplementation on schizophrenia symptoms and association with gastrointestinal functioning: A randomized, placebo-controlled trial. Prim. Care Companion CNS Disord. 16(1):PCC.13m01579.
Donaldson G.P., Ladinsky M.S., Yu K.B., Sanders J.G., Yoo B.B., Chou W.C., Conner M.E., Earl A.M., Knight R., Bjorkman P.J., and Mazmanian S.K. 2018. Gut microbiota utilize immunoglobulin A for mucosal colonization. Science. 360:795–800.
Dong Y. and Benveniste E.N. 2001. Immune function of astrocytes. Glia. 36:180–190.
Dubin P.J. and Kolls J.K. 2008. Th17 cytokines and mucosal immunity. Immunol. Rev. 226:160–171.
Elinav E., Strowig T., Kau A.L., Henao-Mejia J., Thaiss C.A., Booth C.J., Peaper D.R., Bertin J., Eisenbarth S.C., Gordon J.I., and Flavell R.A. 2011. NLRP6 inflammasome regulates colonic microbial ecology and risk for colitis. Cell. 145:745–757.
Emge J.R., Huynh K., Miller E.N., Kaur M., Reardon C., Barrett K.E., and Gareau M.G. 2016. Modulation of the microbiota-gut-brain axis by probiotics in a murine model of inflammatory bowel disease. Am. J. Physiol. Gastrointest. Liver Physiol. 310:G989–G998.
Erny D., de Angelis A.L.H., Jaitin D., Wieghofer P., Staszewski O., David E., Keren-Shaul H., Mahlakoiv T., Jakobshagen K., Buch T., Schwierzeck V., Utermöhlen O., Chun E., Garrett W.S., McCoy K.D., Diefenbach A., Staeheli P., Stecher B., Amit I., and Prinz M. 2015. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 18:965–977.
Farina C., Aloisi F., and Meinl E. 2007. Astrocytes are active players in cerebral innate immunity. Trends Immunol. 28:138–145.
Filipovic B.R. and Filipovic B.F. 2014. Psychiatric comorbidity in the treatment of patients with inflammatory bowel disease. World J. Gastroenterol. 20:3552–3563.
Frost G., Sleeth M.L., Sahuri-Arisoylu M., Lizarbe B., Cerdan S., Brody L., Anastasovska J., Ghourab S., Hankir M., Zhang S., Carling D., Swann J.R., Gibson G., Viardot A., Morrison D., Thomas E.L., and Bell J.D. 2014. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 5:3611.
Fung T.C., Olson C.A., and Hsiao E.Y. 2017. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 20:145–155.
Gaboriau-Routhiau V., Rakotobe S., Lécuyer E., Mulder I., Lan A., Bridonneau C., Rochet V., Pisi A., De Paepe M., Brandi G., Eberl G., Snel J., Kelly D., and Cerf-Bensussan N. 2009. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity. 31:677–689.
Gareau M.G., Wine E., Rodrigues D.M., Cho J.H., Whary M.T., Philpott D.J., MacQueen G., and Sherman P.M. 2011. Bacterial infection causes stress-induced memory dysfunction in mice. Gut. 60:307–317.
Garrett W.S., Lord G.M., Punit S., Lugo-Villarino G., Mazmanian S.K., Ito S., Glickman J.N., and Glimcher L.H. 2007. Communicable ulcerative colitis induced by T-bet deficiency in the innate immune system. Cell. 131:33–45.
Goethel A., Turpin W., Rouquier S., Zanello G., Robertson S.J., Streutker C.J., Philpott D.J., and Croitoru K. 2019. Nod2 influences microbial resilience and susceptibility to colitis following antibiotic exposure. Mucosal. Immunol. 12:720–732.
Golubeva A.V., Joyce S.A., Moloney G., Burokas A., Sherwin E., Arboleya S., Flynn I., Khochanskiy D., Moya-Pérez A., Peterson V., Rea K., Murphy K., Makarova O., Buravkov S., Hyland N.P., Stanton C., Clarke G., Gahan C.G.M., Dinan T.G., and Cryan J.F. 2017. Microbiota-related changes in bile acid & tryptophan metabolism are associated with gastrointestinal dysfunction in a mouse model of autism. EBioMedicine. 24:166–178.
Goshen I., Kreisel T., Ounallah-Saad H., Renbaum P., Zalzstein Y., Ben-Hur T., Levy-Lahad E., and Yirmiya R. 2007. A dual role for interleukin-1 in hippocampal-dependent memory processes. Psychoneuroendocrinology. 32:1106–1115.
Griffin W.S.T., Stanley L.C., Ling C.H.E.N., White L., MacLeod V., Perrot L.J., White C.3., and Araoz C. 1989. Brain interleukin 1 and S-100 immunoreactivity are elevated in Down syndrome and Alzheimer disease. Proc. Natl. Acad. Sci. USA. 86:7611–7615.
Hadden J.W., Hadden E.M., and Middleton E. Jr. 1970. Lymphocyte blast transformation. I. Demonstration of adrenergic receptors in human peripheral lymphocytes. Cell. Immunol. 1:583–595.
Haghikia A., Jörg S., Duscha A., Berg J., Manzel A., Waschbisch A., Hammer A., Lee D.H., May C., Wilck N., Balogh A., Ostermann A.I., Schebb N.H., Akkad D.A., Grohme D.A., Kleinewietfeld M., Kempa S., Thöne J., Demir S., Müller D.N., Gold R., and Linker R.A. 2015. Dietary fatty acids directly impact central nervous system autoimmunity via the small intestine. Immunity. 43:817–829.
Hamada H., Hiroi T., Nishiyama Y., Takahashi H., Masunaga Y., Hachimura S., Kaminogawa S., Takahashi-Iwanaga H., Iwanaga T., Kiyono H., Yamamoto H., and Ishikawa H. 2002. Identification of multiple isolated lymphoid follicles on the antimesenteric wall of the mouse small intestine. J. Immunol. 168:57.
Hansen C.H., Nielsen D.S., Kverka M., Zakostelska Z., Klimesova K., Hudcovic T., Tlaskalova-Hogenova H., and Hansen A.K. 2012. Patterns of early gut colonization shape future immune responses of the host. PLoS ONE. 7:e34043.
Hillman E.T., Lu H., Yao T., and Nakatsu C.H. 2017. Microbial ecology along the gastrointestinal tract. Microbes Environ. 32:300–313.
Hong S., Dissing-Olesen L., and Stevens B. 2016. New insights on the role of microglia in synaptic pruning in health and disease. Curr. Opin. Neurobiol. 36:128–134.
Hooper L.V., Littman D.R., and Macpherson A.J. 2012. Interactions between the microbiota and the immune system. Science. 336:1268–1273.
Hsiao E.Y. 2014. Gastrointestinal issues in autism spectrum disorder. Harvard Rev. Psychiatry. 22:104–111.
Hsiao E.Y., McBride S.W., Hsien S., Sharon G., Hyde E.R., McCue T., Codelli J.A., Chow J., Reisman S.E., Petrosino J.F., Patterson P.H., and Mazmanian S.K. 2013. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell. 155:1451–1463.
Humann J., Mann B., Gao G., Moresco P., Ramahi J., Loh L.N., Farr A., Hu Y., Durick-Eder K., Fillon S.A., Smeyne R.J., and Tuomanen E.I. 2016. Bacterial peptidoglycan traverses the placenta to induce fetal neuroproliferation and aberrant postnatal behavior. Cell Host Microbe. 19:388–399.
Huuskonen J., Suuronen T., Nuutinen T., Kyrylenko S., and Salminen A. 2004. Regulation of microglial inflammatory response by sodium butyrate and short-chain fatty acids. Br. J. Pharmacol. 141:874–880.
Iadecola C. and Anrather J. 2011. The immunology of stroke: From mechanisms to translation. Nat. Med. 17:796–808.
Ivanov I.I., Atarashi K., Manel N., Brodie E.L., Shima T., Karaoz U., Wei D., Goldfarb K.C., Santee C.A., Lynch S.V., Tanoue T., Imaoka A., Itoh K., Takeda K., Umesaki Y., Honda K., and Littman D.R. 2009. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell. 139:485–498.
Jensen C.J., Massie A., and De Keyser J. 2013. Immune players in the CNS: The astrocyte. J. Neuroimmune Pharmacol. 8:824–839.
Karamese M., Aydin H., Sengul E., Gelen V., Sevim C., Ustek D., and Karakus E. 2016. The immunostimulatory effect of lactic acid bacteria in a rat model. Iran. J. Immunol. 13:220–228.
Kawamoto S., Tran T.H., Maruya M., Suzuki K., Doi Y., Tsutsui Y., Kato L.M., and Fagarasan S. 2012. The inhibitory receptor PD-1 regulates IgA selection and bacterial composition in the gut. Science. 336:485–489.
Kawamoto S., Maruya M., Kato L.M., Suda W., Atarashi K., Doi Y., Tsutsui Y., Qin H., Honda K., Okada T., Hattori M., and Fagarasan S. 2014. Foxp3(+) T cells regulate immunoglobulin a selection and facilitate diversification of bacterial species responsible for immune homeostasis. Immunity. 41:152–165.
Kebir H., Kreymborg K., Ifergan I., Dodelet-Devillers A., Cayrol R., Bernard M., Giuliani F., Arbour N., Becher B., and Prat A. 2007. Human TH17 lymphocytes promote blood-brain barrier disruption and central nervous system inflammation. Nat. Med. 13:1173–1175.
Khakh B.S. and Sofroniew M.V. 2015. Diversity of astrocyte functions and phenotypes in neural circuits. Nat. Neurosci. 18:942–952.
Kim M.H., Kang S.G., Park J.H., Yanagisawa M., and Kim C.H. 2013. Short-chain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology. 145:396.
Kim S., Kim H., Yim Y.S., Ha S., Atarashi K., Tan T.G., Longman R.S., Honda K., Littman D.R., Choi G.B., and Huh J.R. 2017. Maternal gut bacteria promote neurodevelopmental abnormalities in mouse offspring. Nature. 549:528–532.
Kimura I., Inoue D., Maeda T., Hara T., Ichimura A., Miyauchi S., Kobayashi M., Hirasawa A., and Tsujimoto G. 2011. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA. 108:8030–8035.
Kinnebrew M.A. and Pamer E.G. 2012. Innate immune signaling in defense against intestinal microbes. Immunol. Rev. 245:113–131.
Kobayashi K.S., Chamaillard M., Ogura Y., Henegariu O., Inohara N., Nuñez G., and Flavell R.A. 2005. Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science. 307:731–734.
Kolb B. and Whishaw I.Q. 1998. Brain plasticity and behavior. Annu. Rev. Psychol. 49:43–64.
Korn T., Anderson A., Bettelli E., and Oukka M. 2007. The dynamics of effector T cells and Foxp3+ regulatory T cells in the promotion and regulation of autoimmune encephalomyelitis. J. Neuroimmunol. 191:51–60.
Kwon O., Lee S., Kim J.H., Kim H., and Lee S.W. 2015. Altered gut microbiota composition in Rag1-deficient mice contributes to modulating homeostasis of hematopoietic stem and progenitor cells. Immune Netw. 15:252–259.
Lakhan S.E. and Kirchgessner A. 2010. Neuroinflammation in inflammatory bowel disease. J. Neuroinflammation. 7:37.
Lee Y.K., Menezes J.S., Umesaki Y., and Mazmanian S.K. 2011. Proinflammatory T-cell responses to gut microbiota promote experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA. 108(Suppl. 1):4615–4622.
Levy O. 2007. Innate immunity of the newborn: Basic mechanisms and clinical correlates. Nat. Rev. Immunol. 7:379–390.
Ley R.E., Hamady M., Lozupone C., Turnbaugh P.J., Ramey R.R., Bircher J.S., Schlegel M.L., Tucker T.A., Schrenzel M.D., Knight R., and Gordon J.I. 2008a. Evolution of mammals and their gut microbes. Science. 320:1647–1651.
Ley R.E., Lozupone C.A., Hamady M., Knight R., and Gordon J.I. 2008b. Worlds within worlds: Evolution of the vertebrate gut microbiota. Nat. Rev. Microbiol. 6:776–788.
Liang S., Wang T., Hu X., Luo J., Li W., Wu X., Duan Y., and Jin F. 2015. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neuroscience. 310:561–577.
Llopis M., Antolin M., Guarner F., Salas A., and Malagelada J.R. 2005. Mucosal colonisation with Lactobacillus casei mitigates barrier injury induced by exposure to trinitronbenzene sulphonic acid. Gut. 54:955–959.
Llopis M., Antolin M., Carol M., Borruel N., Casellas F., Martinez C., Espín-Basany E., Guarner F., and Malagelada J.R. 2009. Lactobacillus casei downregulates commensals’ inflammatory signals in Crohn’s disease mucosa. Inflamm. Bowel Dis. 15:275–283.
Louveau A., Smirnov I., Keyes T.J., Eccles J.D., Rouhani S.J., Peske J.D., Derecki N.C., Castle D., Mandell J.W., Lee K.S., and Harris T.H. 2015. Structural and functional features of central nervous system lymphatic vessels. Nature. 523:337–341.
Luo J., Wang T., Liang S., Hu X., Li W., and Jin F. 2014. Ingestion of Lactobacillus strain reduces anxiety and improves cognitive function in the hyperammonemia rat. Sci. China Life Sci. 57:327–335.
Maes M., Kubera M., Leunis J.C., and Berk M. 2012. Increased IgA and IgM responses against gut commensals in chronic depression: Further evidence for increased bacterial translocation or leaky gut. J. Affect. Disord. 141:55–62.
Mamantopoulos M., Ronchi F., Van Hauwermeiren F., Vieira-Silva S., Yilmaz B., Martens L., Saeys Y., Drexler S.K., Yazdi A.S., Raes J., and Lamkanfi M. 2017. Nlrp6- and ASC-dependent inflammasomes do not shape the commensal gut microbiota composition. Immunity. 47:339–348.
Martin, C.R., and Mayer, E.A. 2017. Gut-brain axis and behavior. In 88th Nestlé Nutrition Institute Workshop, Playa del Carmen, Mexico, September 2016. Vol. 88, pp. 45–53.
Martinez K.A. II, Romano-Keeler J., Zackular J.P., Moore D.J., Brucker R.M., Hooper C., Meng S., Brown N., Mallal S., Reese J., Aronoff D.M., Shin H., Dominguez-Bello M.G., and Weitkamp J.H. 2018. Bacterial DNA is present in the fetal intestine and overlaps with that in the placenta in mice. PLoS ONE. 13:e0197439.
Matcovitch-Natan O., Winter D.R., Giladi A., Aguilar S.V., Spinrad A., Sarrazin S., Ben-Yehuda H., David E., González F.Z., and Perrin P. 2016. Microglia development follows a stepwise program to regulate brain homeostasis. Science. 353:aad8670.
Mayer E.A., Savidge T., and Shulman R.J. 2014. Brain-gut microbiome interactions and functional bowel disorders. Gastroenterology. 146:1500–1512.
Mazmanian S.K., Liu C.H., Tzianabos A.O., and Kasper D.L. 2005. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell. 122:107–118.
McCarthy J., O’Mahony L., O’Callaghan L., Sheil B., Vaughan E.E., Fitzsimons N., Fitzgibbon J., O’Sullivan G.C., Kiely B., Collins J.K., and Shanahan F. 2003. Double blind, placebo controlled trial of two probiotic strains in interleukin 10 knockout mice and mechanistic link with cytokine balance. Gut. 52:975–980.
McFarland L.V., Evans C.T., and Goldstein E.J.C. 2018. Strain-specificity and disease-specificity of probiotic efficacy: A systematic review and meta-analysis. Front. Med. 5:124.
Messaoudi M., Lalonde R., Violle N., Javelot H., Desor D., Nejdi A., Bisson J.F., Rougeot C., Pichelin M., Cazaubiel M., and Cazaubiel J.M. 2011a. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br. J. Nutr. 105:755–764.
Messaoudi M., Violle N., Bisson J.F., Desor D., Javelot H., and Rougeot C. 2011b. Beneficial psychological effects of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in healthy human volunteers. Gut Microbes. 2:256–261.
Milo L.A., Reardon K.A., and Tappenden K.A. 2002. Effects of short-chain fatty acid-supplemented total parenteral nutrition on intestinal pro-inflammatory cytokine abundance. Dig. Dis. Sci. 47:2049–2055.
Mohammadi A.A., Jazayeri S., Khosravi-Darani K., Solati Z., Mohammadpour N., Asemi Z., Adab Z., Djalali M., Tehrani-Doost M., Hosseini M., and Eghtesadi S. 2016. The effects of probiotics on mental health and hypothalamic-pituitary-adrenal axis: A randomized, double-blind, placebo-controlled trial in petrochemical workers. Nutr. Neurosci. 19:387–395.
Mohle L., Mattei D., Heimesaat M.M., Bereswill S., Fischer A., Alutis M., French T., Hambardzumyan D., Matzinger P., Dunay I.R., and Wolf S.A. 2016. Ly6C(hi) monocytes provide a link between antibiotic-induced changes in gut microbiota and adult hippocampal neurogenesis. Cell Rep. 15:1945–1956.
Morelli L. and Capurso L. 2012. FAO/WHO guidelines on probiotics: 10 years later. J. Clin. Gastroenterol. 46:S1–S2.
Nakajima A., Vogelzang A., Maruya M., Miyajima M., Murata M., Son A., Kuwahara T., Tsuruyama T., Yamada S., Matsuura M., and Nakase H. 2018. IgA regulates the composition and metabolic function of gut microbiota by promoting symbiosis between bacteria. J. Exp. Med. 215:2019–2034.
Nayak D., Roth T.L., and McGavern D.B. 2014. Microglia development and function. Annu. Rev. Immunol. 32:367–402.
Ni J., Wu G.D., Albenberg L., and Tomov V.T. 2017. Gut microbiota and IBD: Causation or correlation? Nat. Rev. Gastroenterol. Hepatol. 14:573–584.
Noack H., Lövdén M., and Schmiedek F. 2014. On the validity and generality of transfer effects in cognitive training research. Psychol. Res. 78:773–789.
Ogbonnaya E.S., Clarke G., Shanahan F., Dinan T.G., Cryan J.F., and O’Leary O.F. 2015. Adult hippocampal neurogenesis is regulated by the microbiome. Biol. Psychiatry. 78:e7–e9.
Ohland C.L., Kish L., Bell H., Thiesen A., Hotte N., Pankiv E., and Madsen K.L. 2013. Effects of Lactobacillus helveticus on murine behavior are dependent on diet and genotype and correlate with alterations in the gut microbiome. Psychoneuroendocrinology. 38:1738–1747.
Parfrey L.W., Walters W.A., and Knight R. 2011. Microbial eukaryotes in the human microbiome: Ecology, evolution, and future directions. Front. Microbiol. 2:153–153.
Perez-Muñoz M.E., Arrieta M.-C., Ramer-Tait A.E., and Walter J. 2017. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: Implications for research on the pioneer infant microbiome. Microbiome. 5:48–48.
Pert C.B., Ruff M.R., Weber R.J., and Herkenham M. 1985. Neuropeptides and their receptors: A psychosomatic network. J. Immunol. 135:820s–826s.
PrabhuDas M., Adkins B., Gans H., King C., Levy O., Ramilo O., and Siegrist C.A. 2011. Challenges in infant immunity: Implications for responses to infection and vaccines. Nat. Immunol. 12:189–194.
Prinz M., Erny D., and Hagemeyer N. 2017. Ontogeny and homeostasis of CNS myeloid cells. Nat. Immunol. 18:385–392.
Procaccini C., Pucino V., De Rosa V., Marone G., and Matarese G. 2014. Neuro-endocrine networks controlling immune system in health and disease. Front. Immunol. 5:143.
Proietti M., Cornacchione V., Jost T.R., Romagnani A., Faliti C.E., Perruzza L., Rigoni R., Radaelli E., Caprioli F., Preziuso S., and Brannetti B. 2014. ATP-gated ionotropic P2X7 receptor controls follicular T helper cell numbers in Peyer’s patches to promote host-microbiota mutualism. Immunity. 41:789–801.
Proietti M., Perruzza L., Scribano D., Pellegrini G., D’Antuono R., Strati F., Raffaelli M., Gonzalez S.F., Thelen M., Hardt W.D., and Slack E. 2019. ATP released by intestinal bacteria limits the generation of protective IgA against enteropathogens. Nat. Commun. 10:1–11.
Pusceddu M., Barboza M., Keogh C.E., Schneider M., Stokes P., Sladek J.A., Kim H.J.D., Torres-Fuentes C., Goldfild L.R., Gillis S.E., and Brust-Mascher I. 2019. Nod-like receptors are critical for gut-brain axis signaling in mice. J. Physiol. 597(24):5777–5797.
Rakoff-Nahoum S., Paglino J., Eslami-Varzaneh F., Edberg S., and Medzhitov R. 2004. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell. 118:229–241.
Rao A.V., Bested A.C., Beaulne T.M., Katzman M.A., Iorio C., Berardi J.M., and Logan A.C. 2009. A randomized, double-blind, placebo-controlled pilot study of a probiotic in emotional symptoms of chronic fatigue syndrome. Gut Pathog. 1:6.
Rao R.K. and Samak G. 2013. Protection and restitution of gut barrier by probiotics: Nutritional and clinical implications. Curr. Nutr. Food Sci. 9:99–107.
Rea K., Dinan T.G., and Cryan J.F. 2016. The microbiome: A key regulator of stress and neuroinflammation. Neurobiol. Stress. 4:23–33.
Rhee S.H., Pothoulakis C., and Mayer E.A. 2009. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat. Rev. Gastroenterol. Hepatol. 6:306–314.
Rivera-Chavez F., Zhang L.F., Faber F., Lopez C.A., Byndloss M.X., Olsan E.E., Xu G., Velazquez E.M., Lebrilla C.B., Winter S.E., and Bäumler A.J. 2016. Depletion of butyrate-producing Clostridia from the gut microbiota drives an aerobic luminal expansion of Salmonella. Cell Host Microbe. 19:443–454.
Robertson S.J., Zhou J.Y., Geddes K., Rubino S.J., Cho J.H., Girardin S.E., and Philpott D.J. 2013. Nod1 and Nod2 signaling does not alter the composition of intestinal bacterial communities at homeostasis. Gut Microbes. 4:222–231.
Robertson S.J., Goethel A., Girardin S.E., and Philpott D.J. 2018. Innate immune influences on the gut microbiome: Lessons from mouse models. Trends Immunol. 39:992–1004.
Romijn J.A., Corssmit E.P., Havekes L.M., and Pijl H. 2008. Gut-brain axis. Curr. Opin. Clin. Nutr. Metab. Care. 11:518–521.
Rook G.A., Raison C.L., and Lowry C.A. 2014. Microbiota, immunoregulatory old friends and psychiatric disorders. Adv. Exp. Med. Biol. 817:319–356.
Ross F.M., Allan S.M., Rothwell N.J., and Verkhratsky A. 2003. A dual role for interleukin-1 in LTP in mouse hippocampal slices. J. Neuroimmunol. 144:61–67.
Rothhammer V. and Quintana F.J. 2019. The aryl hydrocarbon receptor: An environmental sensor integrating immune responses in health and disease. Nat. Rev. Immunol. 19:184–197.
Rothhammer V., Mascanfroni I.D., Bunse L., Takenaka M.C., Kenison J.E., Mayo L., Chao C.C., Patel B., Yan R., Blain M., Alvarez J.I., Kébir H., Anandasabapathy N., Izquierdo G., Jung S., Obholzer N., Pochet N., Clish C.B., Prinz M., Prat A., Antel J., and Quintana F.J. 2016. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat. Med. 22:586–597.
Rothhammer V., Borucki D.M., Tjon E.C., Takenaka M.C., Chao C.C., Ardura-Fabregat A., de Lima K.A., Gutiérrez-Vázquez C., Hewson P., Staszewski O., Blain M., Healy L., Neziraj T., Borio M., Wheeler M., Lionel Dragin L., Laplaud D.A., Antel J., Ivan Alvarez J., Prinz M., and Quintana F.J. 2018. Microglial control of astrocytes in response to microbial metabolites. Nature. 557:724–728.
Round J.L., Lee S.M., Li J., Tran G., Jabri B., Chatila T.A., and Mazmanian S.K. 2011. The Toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science. 332:974–977.
Ryu J.H., Kim S.H., Lee H.Y., Bai J.Y., Nam Y.D., Bae J.W., Lee D.G., Shin S.C., Ha E.M., and Lee W.J. 2008. Innate immune homeostasis by the homeobox gene Caudal and commensal-gut mutualism in Drosophila. Science. 319:777–782.
Salvo Romero E., Alonso Cotoner C., Pardo Camacho C., Casado Bedmar M., and Vicario M. 2015. The intestinal barrier function and its involvement in digestive disease. Rev. Esp. Enferm. Dig. 107:686–696.
Sampson T.R. and Mazmanian S.K. 2015. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe. 17:565–576.
Sanctuary M.R., Kain J.N., Chen S.Y., Kalanetra K., Lemay D.G., Rose D.R., Yang H.T., Tancredi D.J., German J.B., Slupsky C.M., and Ashwood P. 2019. Pilot study of probiotic/colostrum supplementation on gut function in children with autism and gastrointestinal symptoms. PLoS ONE. 14:e0210064.
Savignac H.M., Tramullas M., Kiely B., Dinan T.G., and Cryan J.F. 2015. Bifidobacteria modulate cognitive processes in an anxious mouse strain. Behav. Brain Res. 287:59–72.
Schafer D.P. and Stevens B. 2015. Microglia function in central nervous system development and plasticity. Cold Spring Harb. Perspect. Biol. 7:a020545.
Schwab C., Berry D., Rauch I., Rennisch I., Ramesmayer J., Hainzl E., Heider S., Decker T., Kenner L., Müller M., Strobl B., Wagner M., Schleper C., Loy A., and Urich T. 2014. Longitudinal study of murine microbiota activity and interactions with the host during acute inflammation and recovery. ISME J. 8:1101–1114.
Sgritta M., Dooling S.W., Buffington S.A., Momin E.N., Francis M.B., Britton R.A., and Costa-Mattioli M. 2019. Mechanisms underlying microbial-mediated changes in social behavior in mouse models of autism spectrum disorder. Neuron. 101:246–259.
Shanahan M.T., Carroll I.M., Grossniklaus E., White A., von Furstenberg R.J., Barner R., Fodor A.A., Henning S.J., Sartor R.B., and Gulati A.S. 2014. Mouse Paneth cell antimicrobial function is independent of Nod2. Gut. 63:903–910.
Shanks N., Windle R.J., Perks P.A., Harbuz M.S., Jessop D.S., Ingram C.D., and Lightman S.L. 2000. Early-life exposure to endotoxin alters hypothalamic-pituitary-adrenal function and predisposition to inflammation. Proc. Natl. Acad. Sci. USA. 97:5645–5650.
Sharkey K.A. and Mawe G.M. 2002. Neuroimmune and epithelial interactions in intestinal inflammation. Curr. Opin. Pharmacol. 2:669–677.
Shulzhenko N., Morgun A., Hsiao W., Battle M., Yao M., Gavrilova O., Orandle M., Mayer L., Macpherson A.J., McCoy K.D., and Fraser-Liggett C. 2011. Crosstalk between B lymphocytes, microbiota and the intestinal epithelium governs immunity versus metabolism in the gut. Nat. Med. 17:1585–1593.
Siegrist C.A. 2001. Neonatal and early life vaccinology. Vaccine. 19:3331–3346.
Singh P.K., Chopra K., Kuhad A., and Kaur I.P. 2012. Role of Lactobacillus acidophilus loaded floating beads in chronic fatigue syndrome: Behavioral and biochemical evidences. Neurogastroenterol. Motil. 24:366–e170.
Slack E., Hapfelmeier S., Stecher B., Velykoredko Y., Stoel M., Lawson M.A., Geuking M.B., Beutler B., Tedder T.F., Hardt W.D., and Bercik P. 2009. Innate and adaptive immunity cooperate flexibly to maintain host-microbiota mutualism. Science. 325:617–620.
Smith K., McCoy K.D., and Macpherson A.J. 2007. Use of axenic animals in studying the adaptation of mammals to their commensal intestinal microbiota. Semin. Immunol. 19:59–69.
Smith P.A. 2015. The tantalizing links between gut microbes and the brain. Nature. 526:312–314.
Sonnenburg J.L., Angenent L.T., and Gordon J.I. 2004. Getting a grip on things: How do communities of bacterial symbionts become established in our intestine? Nat. Immunol. 5:569–573.
Spulber S., Mateos L., Oprica M., Cedazo-Minguez A., Bartfai T., Winblad B., and Schultzberg M. 2009. Impaired long term memory consolidation in transgenic mice overexpressing the human soluble form of IL-1ra in the brain. J. Neuroimmunol. 208:46–53.
Stanley L.C., Mrak R.E., Woody R.C., Perrot L.J., Zhang S., Marshak D.R., Nelson S.J., and Griffin W.S.T. 1994. Glial cytokines as neuropathogenic factors in HIV infection: Pathogenic similarities to Alzheimer’s disease. J. Neuropathol. Exp. Neurol. 53:231–238.
Steenbergen L., Sellaro R., van Hemert S., Bosch J.A., and Colzato L.S. 2015. A randomized controlled trial to test the effect of multispecies probiotics on cognitive reactivity to sad mood. Brain Behav. Immun. 48:258–264.
Strandwitz P. 2018. Neurotransmitter modulation by the gut microbiota. Brain Res. 1693:128–133.
Theis K.R., Romero R., Winters A.D., Greenberg J.M., Gomez-Lopez N., Alhousseini A., Bieda J., Maymon E., Pacora P., Fettweis J.M., and Buck G.A. 2019. Does the human placenta delivered at term have a microbiota? Results of cultivation, quantitative real-time PCR, 16S rRNA gene sequencing, and metagenomics. Am. J. Obstetr. Gynecol. 220:267.e1–267.e39.
Tremblay M.-È., Stevens B., Sierra A., Wake H., Bessis A., and Nimmerjahn A. 2011. The role of microglia in the healthy brain. J. Neurosci. 31:16064–16069.
Urakubo A., Jarskog L.F., Lieberman J.A., and Gilmore J.H. 2001. Prenatal exposure to maternal infection alters cytokine expression in the placenta, amniotic fluid, and fetal brain. Schizophr. Res. 47:27–36.
Vaghef-Mehrabany E., Alipour B., Homayouni-Rad A., Sharif S.K., Asghari-Jafarabadi M., and Zavvari S. 2014. Probiotic supplementation improves inflammatory status in patients with rheumatoid arthritis. Nutrition. 30:430–435.
Vaishnava S., Behrendt C.L., Ismail A.S., Eckmann L., and Hooper L.V. 2008. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interface. Proc. Natl. Acad. Sci. USA. 105:20858–20863.
van De Sande M.M., van Buul V.J., and Brouns F.J. 2014. Autism and nutrition: The role of the gut-brain axis. Nutr. Res. Rev. 27:199–214.
van Langenberg D.R., Yelland G.W., Robinson S.R., and Gibson P.R. 2017. Cognitive impairment in Crohn’s disease is associated with systemic inflammation, symptom burden and sleep disturbance. United European Gastroenternol. J. 5:579–587.
Vijay-Kumar M., Aitken J.D., Carvalho F.A., Cullender T.C., Mwangi S., Srinivasan S., Sitaraman S.V., Knight R., Ley R.E., and Gewirtz A.T. 2010. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science. 328:228–231.
Vuong H.E. and Hsiao E.Y. 2017. Emerging roles for the gut microbiome in autism spectrum disorder. Biol. Psychiatry. 81:411–423.
Walker L.S., Garber J., Smith C.A., Van Slyke D.A., and Claar R.L. 2001. The relation of daily stressors to somatic and emotional symptoms in children with and without recurrent abdominal pain. J. Consult. Clin. Psychol. 69:85–91.
Wang H., Lee I.S., Braun C., and Enck P. 2016. Effect of probiotics on central nervous system functions in animals and humans: A systematic review. J. Neurogastroenterol. Motil. 22:589–605.
Weltens N., Iven J., Van Oudenhove L., and Kano M. 2018. The gut-brain axis in health neuroscience: Implications for functional gastrointestinal disorders and appetite regulation. Ann. N.Y. Acad. Sci. 1428:129–150.
Williamson L.L., Sholar P.W., Mistry R.S., Smith S.H., and Bilbo S.D. 2011. Microglia and memory: Modulation by early-life infection. J. Neurosci. 31:15511–15521.
Yelin I., Flett K.B., Merakou C., Mehrotra P., Stam J., Snesrud E., Hinkle M., Lesho E., McGann P., McAdam A.J., and Sandora T.J. 2019. Genomic and epidemiological evidence of bacterial transmission from probiotic capsule to blood in ICU patients. Nat. Med. 25:1728–1732.
Yoo B.B. and Mazmanian S.K. 2017. The enteric network: Interactions between the immune and nervous systems of the gut. Immunity. 46:910–926.
Zeng Z., Surewaard B.G.J., Wong C.H.Y., Guettler C., Petri B., Burkhard R., Wyss M., Le Moual H., Devinney R., Thompson G.C., Blackwood J., Joffe A.R., McCoy K.D., Jenne C.N., and Kubes P. 2018. Sex-hormone-driven innate antibodies protect females and infants against EPEC infection. Nat. Immunol. 19:1100–1111.
Zijlmans M.A., Korpela K., Riksen-Walraven J.M., de Vos W.M., and de Weerth C. 2015. Maternal prenatal stress is associated with the infant intestinal microbiota. Psychoneuroendocrinology. 53:233–245.
Zmora N., Zilberman-Schapira G., Suez J., Mor U., Dori-Bachash M., Bashiardes S., Kotler E., Zur M., Regev-Lehavi D., Brik R.B.Z., Federici S., Cohen Y., Linevsky R., Rothschild D., Moor A.E., Ben-Moshe S., Harmelin A., Itzkovitz S., Maharshak N., Shibolet O., Shapiro H., Pevsner-Fischer M., Sharon I., Halpern Z., Segal E., and Elinav E. 2018. Personalized gut mucosal colonization resistance to empiric probiotics is associated with unique host and microbiome features. Cell. 174:1388–1405.
Zonis S., Pechnick R.N., Ljubimov V.A., Mahgerefteh M., Wawrowsky K., Michelsen K.S., and Chesnokova V. 2015. Chronic intestinal inflammation alters hippocampal neurogenesis. J. Neuroinflammat. 12:65.
Information & Authors
Information
Published In

LymphoSign Journal
Volume 7 • Number 1 • March 2020
Pages: 1 - 23
History
Received: 18 November 2019
Accepted: 17 January 2020
Accepted manuscript online: 27 January 2020
Copyright
© 2020.
Authors
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
EloisaSalvo-Romero, PatriciaStokes, and Mélanie G.Gareau. 2020. Microbiota-immune interactions: from gut to brain. LymphoSign Journal.
7(1): 1-23. https://doi.org/10.14785/lymphosign-2019-0018
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item
View Options
View options
Login options
Check if you access through your login credentials or your institution to get full access on this article.
